植物群落
出自 MBA智库百科(https://wiki.mbalib.com/)
植物群落(Plant Community)
目錄 |
什麼是植物群落[1]
植物群落是指在一定地段內,具有一定的植物種類組成、外貌和空間結構,各種植物之間及植物與環境之間彼此影響、相互作用的植物群體組合單元。一個地區全部植物群落共同形成的植物覆被層構成植被,植物群落是植被的結構單元。
植物群落的特征[1]
植物群落具有以下基本特征。
1.具有一定群落外貌
群落外貌即群落的外部形態。由於組成群落的植物種類不同,一個群落中的植物個體,分別處於不同的高度和密度,群落反映到人們眼中的樣子也不同。人們到野外觀察的時候,很容易辨認出來,這是森林、那是草原,就是由於群落的外貌不同。植物群落的外貌通常由構成群落的植物的生活型組成所決定。
2.具有一定群落結構
植物群落是植被的結構單元,具有一系列的結構特點。例如,形態結構上,包括垂直結構與水平結構、成層性、種的分佈格局等。
3.具有一定種類組成
每個植物群落都由一定的植物種類組成。種類組成是區分不同植物群落的首要特征。一個植物群落中種類成分的多少及每種個體的數量,是度量群落多樣性的基礎。
4.形成一定的群落內部環境
植物群落對其居住環境產生重大影響,並形成群落內部特有的環境。例如,森林中的環境與周圍的裸地就有很大的不同,包括光照、溫度、濕度與土壤等都經過了植物群落的改造。即使植物非常稀疏的荒漠群落,土壤等環境條件也有明顯改變。
5.具有一定的動態變化特征
群落是生態系統中有生命的部分。生命的特征是不停地運動。群落也是如此,其運動形式包括波動和演替等。
6.可以對群落進行分類與排序
群落因種類組成、外貌與結構、生態環境、演替特點的不同,可以區分為不同的類型,進行群落的分類。把一個地區內調查的植物群落樣地,按相似度排定各樣地的位序,從而分析各樣地之間及其與周圍生境之間的相互關係,稱為植物群落的排序。
植物群落的性質[1]
1.整體性說(有機體學派、準有機體學派)
在植物生態學發展的早期,美國生態學家克萊門茨(1916;1928)認為,植物群落類似一個有機體:其理論根據是:任何一個植物群落都要經歷一個從先鋒階段到相對穩定的頂極階段的演替過程,如果時間充足的話,森林區的一片沼澤最終會演替為森林植被。這個演替過程類似於一個有機體的生活史。因此,群落像一個有機體一樣,有誕生、生長、成熟和死亡的不同發育階段,就像一個有機體的不同發育時期。他認為這種比擬是特別真實的,每一個頂極群落破壞後都能夠通過基本上是同樣形式的發展階段再到達頂極階段。
著名的法瑞學派創始人Braun-Blanquet(1928;1932)和另外一些學者(Nichols,1917;Warming,1909)把植物群落比擬為物種,把植物群落的分類看作與生物有機體的分類相似。正如物種是生物分類的基本單位一樣,植物群落的最基本分類單位(植物群叢)則是植被的基本單位。在這種比擬中,他忽略了物種是一個遺傳單位,而植物群落卻在遺傳上與其他相似的群落無關。群落間的從屬關係不同於有機體之間的關係,它們僅是建立在某種結構和組成特征相似的基礎上。
英國生態學家坦斯利(1920)雖然認為克萊門茨的有機體觀點不正確,並指出在植物群落中,有些種群是獨立的,它們在別的群落中也能很好地生長髮育,相反,有些種群卻具有強烈的依附性,即只能在這樣的群落中而不能在別的群落中生長,群落中一些種群和某類群落並非嚴密結合,但他仍然主張群落具有整體性,應作為整體來研究,並用準有機體字樣來形容群落,在此基礎上他提出了生態系統概念,強調各個組成成分之間存在緊密聯繫。
以上學者的看法雖然有一定的差別,但都認為,植物群落是一個整體,從整體水平對外界發生反應。在某種程度上,類似於一個有機體,有自己的發生、發展與演變過程,各個群落間存在著明顯的邊界,植物群落可以和植物種一樣進行分類與命名。每個植物群落是一個整體,而植被由不同的群落組成,所以植被具有不連續性。對群落這樣的看法被稱為整體性說。包括兩個方面的基本認識:①群落的整體性(有機性或準有機性);②植被的不連續性。
2.個別性說(連續性觀點、個體學派)
在整體性說流行時,H.A.Gleason經過審慎研究後,在1926年發表了“植物群叢中的個體論概念(individualistic concept)”一文,提出了恰好相反的觀點,認為任何有關群落與有機體比擬都是欠妥的。因為群落的存在依賴於特定的生境與物種的選擇性,但環境條件在空間與時間上都是不斷變化的,因此群落之間不具有明顯的邊界。他指出長距離內連續觀察的森林群落中,各物種及其多度的更替變化是漸變的,即強調植物群落內物種的個別性(或獨立性)。即使在環境相對同質的地區,仍然可以見到植被的連續性變化,因而不存在群落的邊界,而且在自然界沒有任何兩個群落是相同或相互密切關聯的。傳統群落分類實屬人為產物。由於環境變化而引起的群落差異性是連續的。原蘇聯的R.G.Ramensky和美國的R.H.Whittaker均持類似觀點。美國學者Whittaker(1970)的工作結果表明,在絕大多數情況下,不同種群之間的競爭並不產生明顯的邊界,也並不產生分佈相似的種的集群。種群的中心和邊界是沿著環境梯度散佈的。這種觀察結果與Gleason和Ramensky(1930)所主張的“種的個別性原則”相符合。他還認為,沒有兩個種分佈相同,因為每個種都按照本身遺傳的、生理的和生活周期特性,以及與環境條件和其他種的相互作用方式,以各自的方式分佈。並且,由於不同種群沿著一個梯度分佈,其生態位的重疊是疏鬆的,其中心是分散的,因而大多數群落總是沿著環境梯度連續地相互依次排列,而不形成明顯的、可清楚劃分的界限,植被存在著連續性。
強調種的獨立性,認為植物是以種群(物種)為單位對環境作出反應,不存在明顯的、可清楚劃分界限的群落整體,在連續變化的環境梯度條件下,植被具有連續性,這樣的觀點稱為個體學派。個體學派包括以下兩點基本認識:①種的獨立性;②植被的連續性。
根據這種看法,在研究植物群落時,更多地重視種群特征分析,重視隨環境梯度的變化,種群性質出現的變化。強調種的獨立性。認為植物群落可以出於某種需要進行分類,但其邊界是主觀人為劃定的,沒有客觀性。他們用梯度分析與排序等定量方法研究植被,證明群落並不是一個個分離的、有明顯邊界的實體,多數情況下是在空間和時間上連續的一個系列。
植物群落的分類[1]
一、植物群落分類的原則
按照傳統的整體學派觀點,可以對植物群落進行分類。不同國家植被條件不同,植被研究中,對植物群落分類採用的分類標準也有所差異。目前我國採用的是“植物群落學~生態學原則”,它是以植物群落本身特征作為分類依據,同時又考慮到群落的生態關係。分類依據包括以下幾方面。
(1)植物種類組成
種類組成是群落的最主要特征。群落的其他特征幾乎全由這一特征決定。因此,在進行植物群落分類時考慮群落的種類組成是很自然的。中國植被主要採用優勢種原則。把群落中各個層或層片中數量最多、蓋度最大、對群落作用最明顯的種,稱為優勢種。在主要層片(優勢層片)中的優勢種稱建群種。在植被類型複雜,採用優勢種原則有困難時,則可考慮採用生態幅度狹窄,對該類型有標誌作用的標誌種作為劃分類型的標準。
(2)外貌和結構
外貌和結構是植物群落分類的傳統依據。對未經充分研究的巨集觀區域的植被進行分類時,採用外貌和結構原則常可得到比較滿意的結果。外貌主要根據生活型來劃分。例如,按照生長型分類系統,從形態學的角度,把植物分為喬木、灌木、草本和葉狀體等類群,以下再按形態和發育節律(落葉、常綠)劃分第二級、第三級等。
(3)生態地理特征
由於歷史原因,有時生活型和外貌不一定完全反映現代環境條件,按外貌原則劃分的植被類型,常常包括了異質的類群。例如,針葉林在生活型和外貌上是相似的,但卻分佈在不同的生境里。在分類時,就應該考慮生態地理的因素,根據分佈地區氣候條件的不同,劃分為寒溫性針葉林、溫性針葉林、暖溫性針葉林、熱性針葉林等。
(4)動態特征
優勢種的原則著重於群落的現狀,沒有特別分出原生類型和次生類型。但是在某些情況下,需要考慮群落的動態演替特征。例如,我國森林區荒山坡灌草叢,按生態外貌原則應該是一獨立的植被類型,但考慮到它的次生性質及與灌叢的演替關係,而把它與灌叢合為同一植被型組。
二、植物群落的分類系統
不同國家、不同地區的植被有各自的特點,不同學派考慮到所在地區的植被特征,往往採用不同的分類原則與依據。因此,形成了不同的植物群落分類系統,代表性的如英美學派、北歐學派、法瑞學派、蘇聯學派等。中國植被分類系統受蘇聯學派的影響較大。在《中國植被》(1980)一書中的系統採用植被型、群系、群叢3個基本分類單位,在各基本分類單位之上,各設一輔助單位,即植被型組、群系組與群叢組。並根據需要,在主分類單位之下設立亞級輔助單位,如植被亞型、亞群系等,以作為該級分類單位的補充,這樣,得到九級分類單位,即植被型組、植被型、植被亞型、群系組、群系、亞群系、群叢組、群叢、亞群叢。
1.植被型組
植被型組是最高級的分類單位。凡建群種生活型相似,群落外貌、形態相似的植物群落聯合為植被型組。我國1980年版《中國植被》共分了10個植被型組:針葉林、闊葉林、灌叢和灌草叢、草原和稀樹幹草原、荒漠、凍原、高山稀疏植被、草甸、沼澤、水生植被。它們主要是根據群落的外貌進行劃分的。
2.植被型
植被型是最重要的高級分類單位,是在植被型組內,建群種生活型一級、二級相同或相近,同時對水熱條件生態關係一致的植物群落聯合。我國植被共分為29個植被型。例如,在針葉林植被型組下有5個植被型:寒溫性針葉林,如興安落葉鬆林;溫性針葉林,如華山鬆林、油鬆林;溫陛針闊葉混交林;暖性針葉林,如馬尾鬆林;熱性針葉林,如臺灣鬆林。
建群種生活型相同或近似,反映了群落進化過程中對環境條件適應途徑的一致,即其生態幅度和適應範圍一致。就地帶性植被而言,植被型是一定氣候區域的產物,就非地帶性植被而言,它是一定的特殊生境的產物。據此確定的植被型,大致有相似的結構、相似的生態性質,以及相似的發生和發展歷史。
3.植被亞型
植被亞型為植被型的輔助單位,在植被型內根據優勢層片或指示層片的差異進一步劃分亞型。這種層片結構的差異,一般是由氣候亞帶的差異或一定的地貌、基質條件差異引起的。例如,寒溫性針葉林植被型下劃分兩個亞型:寒溫性落葉針葉林、寒溫性常綠針葉林。雨林植被型(屬於闊葉林植被型組)下劃分為3個亞型:濕潤雨林、季節雨林、山地雨林。
4.群系組
群系組是在植被型或植被亞型範圍內,根據建群種親緣關係近似(同屬或相近屬)、生活型(三級或四級)近似或生境相近而劃分為群系組。劃入為同一群系組的各群系,其生態特點一定是相似的。例如,典型草原可分出叢生禾草草原、根莖禾草草原和小半灌木草原群系組;溫陛常綠針葉林(植被亞型)可以分出溫陛鬆林、側柏林、鐵杉林等群系組。
5.群系
群系是最重要的中級分類單位。凡是建群種或共建種相同的植物群落聯合為群系,以建群種命名,如馬尾鬆林群系、興安落葉鬆林群系、羊草草原群系。由於建群種或共建種相同,一個群系的結構、區系組成、生物生產力及動態特點都是相似的,因此,採用建群種(共建種)這一指標,就概括了群落各方面的特征。一般情況下,地帶性群系主要分佈在氣候亞帶範圍內,非地帶性群系則局限在某一特定生態因數的一定梯度範圍內。在類型等級上,群系還常局限在某一植被亞型內。但對少數廣生態幅的建群種,也會遇到例外的情況。例如,馬尾鬆林,從北亞熱帶一直分佈到南亞熱帶,同時在幾個氣候亞帶內出現。又如,羊草草原和溝葉羊茅草原,在草甸草原和典型草原內均可遇到。還有極個別的種,如蘆葦,不但在地域上分佈很廣,而且可形成沼澤、鹽生草甸等不同植被類型,但在不同的生境中,其體態、群落外貌和結構均有很大變異。因此,按其最適生境將蘆葦放入沼澤生境。
6.亞群系
亞群系是群系的輔助單位。在生態幅度比較廣的群系內,根據次優勢層片及其所反應的生境條件的差異而劃分為亞群系。例如,羊草草原群系可劃分出羊草一中生雜草、羊草一叢生禾草、羊草一鹽中生雜草3個亞群系。但對大多數群系來說,並不需要劃分亞群系。
7.群叢組
將層片結構相似,而且優勢層片與次優勢層片的優勢種或共優種(某種情況下為標誌種)相同的植物群落聯合為群叢組,如羊草一叢生禾草亞群系之下的羊草一大針茅草原群叢組。
8.群叢
群叢是植被分類的主要基本單位。層片結構相同且各層片優勢種或共優種相同的植物群落聯合為群叢。換言之,屬於同一群叢的植物群落,應具有相同的結構、每層具共同的優勢種類、相同的生態特征、相同的動態特點、相似的生境、相同的季相變化並處於相同的演替階段。命名:列出各層的優勢種,不同層之間用“-”分隔,同層共優種之間用“+”分隔,如馬尾松-杜鵑-鐵芒萁群叢;馬尾松+細葉青岡-杜鵑-鐵芒萁+里白群叢。
植物群落的結構[1]
群落的結構,包括垂直結構和水平結構。
一、植物群落的垂直結構
群落的垂直結構,主要指群落的垂直分層現象,包括層與層片的分化。
1.層的分化
層(1ayer)的分化即不同的生物種出現在地面以上的不同高度和地面以下的不同深度,在整個群落內的垂直高度上出現許多層次的成層現象。
例如,森林植物群落根據組成植物高度的不同,地上部分一般可分為喬木層、灌木層、草本層和地被層4層。由於喬木層扎根很深,灌木及草本扎根較淺,因此,在群落的地下也有成層分化,這種分化和地上的層次分化是相對應的。
這種成層結構使植物充分利用環境中的空間:陽光、水分和礦質營養,擴大了植物利用環境的範圍和效率。例如,在發育成熟的森林中,上層喬木可以充分利用陽光,穿過喬木層的光,有時僅占到達樹冠的全光照的1/10,但林下灌木層卻能利用這些微弱的、並且光譜組成已被改變了的光。在灌木層下的草本層能夠利用更微弱的光,草本層往下還有更耐陰的苔蘚層。
群落對光能的利用效率可以用葉面積指數來衡量。葉面積指數(LAI)是指單位土地面積上植物葉片的單面總面積。LAI=總葉片錶面積/單位土地面積。
一些主要的天然植被類型,其葉面積指數如表1所示,由表1可知,葉面積指數與該群落的光能利用率有直接關係。葉面積指數越大,葉片交錯重疊程度越大。群落對有效光能的利用效率越高。
表1 主要植被類型的葉面積指數與光能利用率(引自Barbour,et a1.1980)
| 植被類型 | LAI | 光能利用率% |
|---|---|---|
| 熱帶雨林 | 10-11 | 1.5 |
| 落葉闊葉林 | 5-8 | 1.0 |
| 北方針葉林 | 9-11 | 0.75 |
| 草原 | 5-8 | 0.50 |
| 凍原 | 1-2 | 0.25 |
| 草原化荒漠 | 1 | 0.04 |
| 農作物 | 3-5 | 0.60 |
註:光能利用率計算方法:單位面積凈生產量(每克物質17.6kJ計)與單位面積全年接收的有效光合輻射(kJ)之比。
森林中的藤本和附生、寄生植物,它們攀援或附著在直立植株的不同高度,往往在整個群落的垂直高度內都有分佈,因而並不形成一個層次,稱為層間植物(interstratumplant)或層外植物(extrastratum plant)。
2.層片的分化
層片(synusia)詞系瑞典植物學家Gains(1918)首創。他將層片劃分為3級:第一級層片是同種個體的組合,第二級層片是同一生活型的不同植物的組合,第三級層片是不同生活型的不同種類植物的組合,即H.Gams的第一級層片指的是種群,第三級層片指的是植物群落。現在群落學研究中一般使用的層片概念,相當於Gams(1918)的第二級層片,即每一個層片均由同一生活型的不同植物所構成。因此,與層的分化只考慮高度不同,層片的分化主要考慮的是植物的生活型。
層片是植物群落結構的一個基本單位,由相同生活型或相似生態要求的種組成,同一層的植物可能屬於不同的層片。例如,針闊葉混交林的喬木層,在層上可能是一層,但在二級層片上,包括了針葉層片和落葉闊葉層片兩個層片。同一層片的植物也可能屬於不同的層。例如,高位芽植物作為一個層片,植物的高度各不相同,可能屬於喬木層或灌木層。
二、植物群落的水平結構
植物群落水平結構的主要特點是種間鑲嵌性,即在植物群落內,種群與種群間分佈的水平結構特征主要表現為“鑲嵌性”,即同種植物常呈斑塊狀聚生在一起,不同物種的斑塊鑲嵌存在。例如,林冠下層光照強度強的林窗處,聚生著陽生灌木及草本,而光照強度弱的地方,只能生長耐陰植物。鑲嵌性主要是由“生態因數的不均勻性”造成的,如小地形的差異、光照的強弱、土壤性質的不同及植物本身的繁殖特性等。
植物群落的種類組成[1]
種類組成是決定群落性質最重要的因素,也是鑒別不同群落類型的基本特征。怎樣知道一個群落含有哪些物種及物種數量的多少呢?對於植物群落來說,通常採用野外實地調查的方法。每類群落所占的空間和位置不同,或大或小,或連續或分散。一般情況下不可能,也不需要把所有地段全面地進行清查,特別是數量特征更不可能如數查清。所以一般採取抽樣調查方法完成這一任務。它的實質是選擇調查有代表性的一定數量的小面積地段進行詳細調查,以此估計推斷此類群落整體。這些小面積地段稱為樣地。每塊樣地應該設在同樣生境下同一類型植物群落中。
1.樣地的形狀
大多採用方形(長方形),故又稱樣方。也有使用圓形樣地的,稱為樣圓。因為樣圓很難划出邊界,不及樣方設置便利,故只在少數情況下使用小樣圓調查,如對草原群落或森林下層草本層調查時可採用。長方形樣地的長軸以平行於等高線為好,否則高差過大,樣地內可能出現生境的變化,不好觀察群落特征。樣方的缺點是周邊長,地處過界的植物增多,取捨不便;直角常測不准,出現閉合差,有時影響調查精度。調查林木蓄積量和草地產草量時要註意這類誤差值。
小型樣方(1m2)用於調查草本群落或者林下草本植物層。大型樣方用於喬木群落或結構稀疏的群落。森林調查中樣方常沿著預定的測線方向呈菱形設置。由中心點向前後定距離拉出樣方對角線長度的4個點,然後從這4個點分別拉直長度恰為樣方邊長的測繩,使其在每一側都恰好交接,就是樣方的邊界,這樣可以避免確定直角的麻煩和誤差。
如果山地坡度陡峻,按上述方法所設樣地的投影面積將與實際面積有誤差,須進行坡度訂正,表2中列有在各種坡度上應採用的一半對角線的長度值。
2.樣地大小
群落的最小面積是指包含群落中大多數植物種類,並能表現群落主要結構特征的最小地段面積,它是進行群落野外調查的最小面積。找出群落最小面積一般使用種一面積曲線法,其方法是:從較小面積開始統計植物種類數目,然後逐次向外圍擴大,同時登記新發現的植物種類,直到基本不再增加新種類為止,最後繪製植物種數的面積曲線:以面積大小為橫軸,以種數為縱軸,填入逐次調查數值,並連成平滑曲線,在曲線由陡變緩轉折相對應的面積就是群落最小面積(圖1)。
下列樣地面積的經驗值可供參考使用。
草本群落:1~1Om2
灌叢:16~1OOm2
單純針葉林:1OOm2(10m×1Om)
復層針葉林、夏綠闊葉林:400~500m2(20m×20m,20m×25m)
亞熱帶常綠闊葉林:800~1200m2
熱帶雨林和山地雨林:2500~4000m2
通過各種群落類型的對比,可以發現,環境條件越優越,組成群落的植物種類就越多,群落的結構就越複雜,群落的最小面積相應地就越大。例如,我國西雙版納南部的熱帶雨林群落,它的最小面積至少為2500m2,其中包含了組成群落的主要高等植物130種左右;而東北小興安嶺紅鬆林的一個群落中,最小面積約為400m2,包含主要的高等植物40種左右。
3.樣地數目
最小面積是群落結構的一項特征,也是設定樣地面積的依據。但一塊“最小面積”的樣地並不能包含一個群落的全部植物種類,更不要說所屬群落類型的全部植物種類了。確定一個群落的特征,需要有多個群落樣地資料,否則就不完整。
樣地數目的多少,取決於群落結構的複雜程度。如果群落內部分佈和結構都較均一,少數樣地就能很好地表現出群落特征。當群落的結構複雜並且變化較大,植物分佈不規則時,必須增加樣地數目才能提高調查資料的可靠性。但調查樣地是要付出人力、物力的,所以要考慮經濟效益。究竟取多少樣方,這決定於研究的精度要求。根據統計檢驗理論,多於30個數值(大樣本)才較可靠。
此外,所有樣地必須依照順序編號,或再加以分類排號,這樣做能避免混亂,整理資料時用代號也很方便。
4.樣地的佈局
一塊樣地的面積可能僅占某類群落在一個地區總面積幾百分之一或幾千分之一,各個樣地在這一範圍內應當怎樣佈局也很重要,它影響著調查結果的準確程度。
(1)主觀(有意識)取樣
就是從群落所在空間中,主觀地選擇被認為有代表性的(典型的)或者有某種特點的地塊作為調查樣地,這在路線式粗放調查中常常採用。所選樣地要註意環境條件的一致性與群落外貌的一致性,最好處於群落的中心部位,避免過渡地段。但這種數據容易有偏差或遺漏,特別是一些不顯眼的、較小或分散的特征可能被忽視,因此,這種數量資料不能用於統計分析。
(2)系統取樣(機械取樣)
就是嚴格按照一定的規則(方向和距離)確定樣地位置。在面積十分廣闊的森林和草地中這是一種通用方法。它的優點是布點均勻,定址簡便。缺點是當群落的某些特征呈不規則的隨機分佈時,就可能使某些特征不能如實表露;或者某些呈規律分佈的特征(如沙丘和丘間窪地)恰好和樣地分佈規律不一致,其結果可能就不能反映規律性。
(3)隨機抽樣
這種抽樣的基本原則是,使某類群落各個部分都有同等機會(等概率)被抽取做樣地。為此,應當把要調查的地段分成大小均勻的若幹部分,每部分都編號或確定坐標位置,利用隨機數字表、擲骰、抽簽等方法隨機選出一定數量的、占有一定位置的樣地。地塊的選擇可直接在現場進行,也可選在地圖上進行,然後在現場落實。此法優點是,調查的數據可以應用於各種統計處理,進行可靠性檢驗。許多人認為這是唯一科學有效的方法,但缺點是它常需要較多的樣地,工作量較大,也有個別的種類易被遺漏。
(4)部分隨機取樣(分層抽樣)
部分隨機取樣為系統取樣和隨機取樣的混合法。先將群落地段系統均勻劃分好地段(編號),每個分地段中再隨機抽取樣地。這使所研究的地段內各點被抽取的機會加大了,資料也能用於統計,但是工作量仍然較大。
經過樣地調查,可以發現,有一些群落,所含有的物種數量較多,有些則較少。有些群落的結構較為複雜,有些較為簡單。
一般來說,環境條件越優越,群落的結構越複雜,組成群落的生物種類越多;環境條件越惡劣,群落的結構越簡單,組成群落的生物種類就越少。
群落的複雜程度和物種多少可以用物種多樣性來表述。
物種多樣性(species diversity)是生物多樣性的一個方面,是指一個群落中物種數目的多少及物種個體數目分佈的均勻程度,表示物種的豐富度及其可持續程度。包括兩個方面的內容。
1)種的豐富度(richness of species):物種數量的多少。
2)物種分佈的均勻程度(eveness of species distribution):個體數量在種間分佈的均勻程度。
物種多樣性表達的是物種豐富度及其可維持程度。例如,假設共有100個個體,有A和B兩個物種,甲地有A與B各50個個體,乙地A有99個個體,B有1個個體,則甲地的可維持度就遠高於乙地。因為乙地B物種的1個個體如果死亡,則乙地就只剩A這1個物種了。
物種多樣性指數表示方法很多。以Simpson(辛普生)多樣性指數和Shannon-Wiener指數(香農一威納指數)最為常用。
辛普生多樣性指數(Simpson’s diversity index)的生物學意義是:在很大的群落中,隨機抽取兩個個體,它們為不同物種的概率是多少?Simpson於1949年提出:在不限大小的群落中,隨機抽取得到兩個個體,它們是同一物種的概率與物種多樣性有關。例如,在加拿大北部寒帶森林中,隨機採取兩株樹它們屬於同一個種的概率就很高,相反,如在熱帶雨林中隨機取樣,兩株樹是同一種的概率很低。他從這個想法出發得出多樣性指數。
辛普生多樣性指數=隨機取樣的兩個個體屬於不同種的概率
=1-隨機取樣的兩個個體屬於同種的概率
設總個體數為N,第i個物種的個體數為ni,Pi = ni / N(Pi為相對多度;除此之外,也可用相對優勢度、相對蓋度等)。辛普生多樣性指數D的計算公式為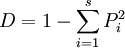
例如:總個體數為N,第i個物種的個體數為ni,Pi = ni / N;
假設N=1000,共2個物種,ni = 500,n2 = 500,則:
P1 = 1 / 2,P2 = 1 / 2,L = 1 − [(P1)2 + (P2)2] = 0.5。
假設ni = 1,ni = 990,則:P1 = 1 / 1000,P2 = 999 / 1000,L = 1 − [(P1)2 + (P2)2] = 0。
辛普生多樣性指數的生物學意義表示的是在群落中任意選取兩個個體,這兩個個體是不同物種的概率。其值大小為0-1。辛普生指數值越大,則表示該群落的物種多樣性越大。
Shannon-Wiener指數(香農-威納指數)的計算公式為
式中,H為Shannon-Wiener指數,與Simpson指數相同;Pi = ni / N,Pi為第i種的相對多度;Ni為第i種的個體數;N為群落中的個體總數;S為物種種數。N也可以採用其他數量參數,如重要值、密度、頻度等。Shannon-Wiener指數來源於信息理論。它的計算公式表明,群落中生物種類增多代表了群落的複雜程度增高,即H值越大,群落所含的信息量越大,群落的物種多樣性越大。
群落物種多樣性越大,則含有的生物基因越豐富,食物鏈越複雜,群落的自我調節能力就越強,生態平衡就越不容易遭到破壞,這叫做物種多樣性導致群落穩定性原則。
此外,在群落交錯區(ecotone),即兩個或多個群落之間的過渡地帶,如海陸交界的潮間帶、河口灣、森林與草原等交界地帶的群落中,生物種類和數量常比單一類型群落中的多,物種多樣性指數高,這種現象稱為邊緣效應(edge effect)。
群落交錯區,又稱生態交錯區或生態過渡帶,如森林和草原之間的森林草原地帶,兩個不同的森林類型之間或兩個草本群落之間的地帶等。這種過渡帶有的寬、有的窄;有的是逐漸變化,有的突然變化。群落交錯區是一個交叉地帶,也是一個種群競爭的緊張地帶,往往包括包含兩個重疊群落中的一些種及交錯區本身所特有的種,這是因為群落交錯區的環境條件比較複雜,能為不同生態類型的植物定居,並且為更多的動物提供食物、營巢和隱蔽條件。例如,我國大興安嶺森林邊緣,每平方米植物達30種以上,明顯高於其內側的森林群落與外側的草原群落。
群落成員型是根據物種在群落中的作用和地位而劃分的物種類群。假設常綠闊葉林樣方,400m2以內,喬木層內大頭茶共100株,栲樹10株,馬尾松3株,那麼明顯可以看出,同樣是該群落內的組成物種,大頭茶在群落中所占地位最重要,其他物種則較次要。因此,可以區分出優勢種、伴生種等不同的類型。
優勢種:群落的優勢種類,是指群落每層中個體數量大、生物量大、覆蓋程度大的生物種類。
建群種群落的主要層次(優勢層)的優勢種稱為建群種,它決定著群落的內部結構和特殊環境條件。
共優種:群落各層的優勢種如果不止一種,則稱為共優種。
共建種:如果群落中有不止一個的建群種則稱為共建種,即在優勢層中有兩個以上的種占優勢。
如果群落中的建群種只有一個,稱為“單優種群落”。如果具有兩個或兩個以上同等重要的建群種,就稱為“共優種群落”或“共建種群落”。熱帶森林幾乎全是共建種群落,北方森林和草原則多為單優種群落,但有時也存在共優種,如由貝加爾針茅和羊草共建的草甸植物群落。
生態學上的優勢種對整個群落具有控制性影響。如果把優勢種去除,必然導致群落性質和環境的變化;但若把非優勢種去除,只會發生較小的或不顯著的變化。因此,不僅要保護那些珍稀瀕危植物,而且也要保護那些建群植物和優勢植物,它們對生態系統的穩定起著舉足輕重的作用。
附屬種:這類植物雖然在群落中出現,參加到群落的組成中去,但對群落內環境所發生的作用不及優勢種,即群落中除優勢種以外的其他生物種類。
附屬種中,根據作用不同,又區分為以下幾種。
亞優勢種:個體數量和作用都次於優勢種,但在決定群落性質和控制群落環境方面仍起著一定作用的植物種。
伴生種:群落中相當穩定的與優勢種伴生在一起的種類。雖然在群落中出現,但對群落內環境所起的作用不及優勢種和亞優勢種。
標誌種:生態幅度狹窄對群落有標誌作用的物種。
偶遇種:在群落中出現頻率很低的種類。可能是由於外界因素(如動物、人類作用)的影響偶然進入群落的物種,也可能是衰退中的殘遺種。
同一個植物種在不同的群落中可以以不同的群落成員型出現。例如,在內蒙古高原中部排水良好的壤質慄鈣土上,針茅是建群種,羊草則是亞優勢種或伴生種;但在地形略為低凹,有地表徑流補給的地方,羊草則是建群種,針茅退居其次。而當強度放牧時,小半灌木冷蒿成為建群種,羊草和針茅成為次要成分。
1.多度
多度(abundance)表示一個種在群落中的個體數量。植物群落中植物種間的個體數量對比關係,可以通過各個種的多度來確定。多度的統計方法,通常有兩種:一種是個體的直接計演算法,即“記名計演算法”,即在一定面積的樣地中,直接點數各種群的個體數目。在森林群落對喬木層的調查中,或者詳細的群落研究中常用;另一種是目測估計法。一般在植物個體數量多而植物體形小的群落(如灌木、草本群落),或者在踏察中常用。目測估計法是按預先確定的多度等級來估計單位面積上個體的多少。
常用的多度等級是德氏(Drude)的七級多度制:Soc.極多;Cop3.很多;Cop2.多;Cop1.尚多;Sp.少;S01.稀少;Un.單一個體。
相對多度(relative abundance):樣地內某一種的個體數占全部種個體數的百分比。
RA=N(某個種個體數)/∑N(全部種個體數之和)×100%
2.密度
密度(density)是指單位面積上的植物株數,用公式表示為
d=N/S
式中,d為密度;N為樣地內某種植物的個體數目;S為樣地面積。
密度的倒數即每株植物所占的單位面積。
樣地內某一種的密度占全部種密度總和的百分比稱為相對密度(relative density)。相對密度:RD=d/∑d×100%。在計算上,等於相對多度。
3.蓋度
蓋度分為以下幾種。
投影蓋度:植物地上部分垂直投影所覆蓋的面積,簡稱蓋度。
郁閉度:森林上層總投影覆蓋地面的比率。大小值用0~1來表示。0代表覆蓋度為0,1代表覆蓋度為100%。
基蓋度:植物基部覆蓋面積。喬木通常用離地1.3m高處的胸高斷面積作為基蓋度。草本以離地面1英寸(2.54cm)高度的斷面積計算。
顯著度(prominance):樣地中一個喬木種的所有個體基蓋度之和,也稱優勢度(dominant)。
相對顯著度(relative pominance):群落中某一物種的顯著度占所有種顯著度之和的百分比。RP=P/∑P×100%
4.頻度
頻度(frequency)是指某物種在調查樣地內出現的頻率。
頻度一某物種出現的樣地數/樣地總數×100%
相對頻度:某一物種的頻度與全部物種頻度之和的比。
RF=F(某個種頻度)/∑F(全部種頻度之和)×100%
丹麥學者拉恩基爾(C.Raunkiaer)用0.1m2的小樣圓任意投擲,將小樣圓內的所有植物種類加以記載,就得到每個小樣圓的植物名錄,然後計算每種植物出現的次數與樣圓總數之比,得到歐洲草地群落中各個種的頻度,並根據8000多種植物的頻度統計結果,編製出標準頻度圖解。通過樣圓對草原植物群落進行數量統計,是一種方便、常用的方法。
5.重要值
重要值(important value)是美國威斯康星(Wisconsin)學派提出的用來表示某個種在群落中地位和作用的綜合數量指標。因為它簡單、明確,所以得到了廣泛應用。
主要用於森林群落喬木層個體重要值的計算。
重要值=相對密度(%)+相對頻度(%)+相對顯著度(%)
IV=RD%+RF%+RP%
結果值常再除以300或3。重要值最大的種即為優勢種。
6.高度
高度為測量植物體體長的一個指標。測量植物取其自然高度或絕對高度:高度分為最大高度、最小高度、平均高度、枝下高度等,要分別測定。在森林群落中,可以使用測高儀測定喬木的高度。
7.體積
體積是生物所占空間大小的度量。在森林植被研究中,這一指標特別重要。在森林經營中,通過體積的計算可以獲得木材生產量(材積)信息。木材的體積稱為材積,可以通過下式計算:材積=胸高斷面積×樹高×形數
形數是樹幹體積與等高同底的圓柱體體積之比。這是由於樹木體形是圓錐形而不是圓柱形,而圓錐形的尖削度又因各個樹種不同,因此在斷面積乘樹高獲得圓柱體體積之後,必須按不同樹種乘以該樹種的形數(在制就的森林調查表中查得),才能獲得喬木的體積。草本植物或小灌木體積的測定,可用排水法進行。
8.重量
重量(質量)是用來衡量植物種群生物量(biomass)或現存量(standing crop)多少的指標。生物量指某一時刻單位面積內實際存活的有機物質總量,通常用kg/m2或t/hm2表示。廣義的生物量是生物在某一特定時刻單位空間的個體數、重量或其含的能量,可用於指某種群、某類群或整個生物群落的生物量。狹義的生物量僅指重量,可以是鮮重或乾重。某一特定時刻的生物量稱為現存量(standing crop)。生產力則是某一時間段內由活的生物體新生產出的有機物質總量。在草原植被研究中,重量指標特別重要。單位面積或容積內某一物種的重量占全部物種總重量的百分比稱為相對重量。
植物群落的內部環境[1]
環境既是形成植物群落的要素,也是群落存在的條件。反過來,群落的存在又必然對環境起到影響,在一定程度上改變環境。一方面,群落的組成、結構、功能、成因、動態、分佈無不受到環境條件的制約;另一方面,群落生活也影響著環境,決定著環境的許多特性,並創造著群落的內部環境。在植物群落內部,由於群落本身的作用而對環境產生影響,在群落內會形成特殊的生境,稱為植物群落的內部環境。以森林為例,在森林植物群落內,與森林外相比,生境有如下變化:弱光、並且主要為漫射光;弱風至靜風;溫差小;濕度大;土壤腐殖質多;氧氣濃度高,CO2濃度較小;且CO2濃度一般從近地面處至林冠逐漸減少等。
一、群落內的光照
在群落內部,由於植物群落對光輻射的吸收、反射和透射作用,所以無論是在森林還是草本群落的植冠下,入射的光量總比其他條件相同而沒有植被的陸地錶面少得多。
穿過枝葉間隙射入群落內部的直射光,形成大小不等的光斑,雖然其成分改變很少,但照射強度顯著減弱。
通過群落上層進入林下的光,由於群落對光的“過濾”作用,光的成分和強度都發生了變化。由於大部分有效輻射被葉片吸收,林下以散射光為主,強度上也顯著降低。
可使用光照度計,通過測定相對光照度來定量光減弱的程度。
二、群落內的溫度
群落內與群落外的溫度相比波動較為緩和。一般群落內的平均溫度白天和夏天低於群落外空曠地面,而夜間和冬季則高於群落外空曠地面。晝夜和全年的溫度變幅較小。
群落的結構越複雜,郁閉度越高,對溫度的作用越顯著。大面積的森林,群落對溫度的作用不僅在林內,還可以影響一定範圍的地方氣候。
三、群落內的水分
植物群落能截留降水,保蓄水分。
如在河流的上游營造大面積森林,可以起到涵養水源的作用;在山坡地帶建造森林,可以減少地面徑流,防止水土流失。因此,植物群落在涵養水源、保持水土方面有重要意義。
降落到群落的水分,一部分到達地面,一部分被樹冠截留。群落截留水分的能力,一與群落結構、郁閉度有關;二與降水強度有關,降水強度越小,截留越多;三與土壤的結構和性質有關。土壤結構差、質地粗,保水能力差;四與地形起伏大小有關,地形起伏大也不能有效地保留水分。
四、群落內的空氣
植物群落對空氣的改變主要表現在兩個方面。
空氣成分發生變化:由於植物的光合作用,群落內CO2濃度降低,O2濃度增大,尤其是靠近林冠上層,這種變化最為明顯。
森林不但可以增加氧氣,還可以吸附粉塵;對一些有毒氣體如SO2、HF、Cl2等有一定吸收能力,是天然的空氣凈化器。
抑制風速:由於群落對氣流的阻抑,群落內表現為弱風,在結構複雜郁閉度高的森林內,甚至表現為靜風狀態。
五、群落內的土壤
植物群落對土壤最明顯的作用是可以增加土壤腐殖質含量,增強土壤肥力。
此外,植物群落在改善土壤理化性質、優化土壤結構、固定流沙、防止和治理鹽鹼化方面都有顯著作用。
植被類型通常與一定的土壤條件密切關聯,如熱帶雨林—磚紅壤、北方針葉林—灰化土、溫帶草原—黑鈣土或慄鈣土等,二者共同受到氣候條件的控制。
綜上所述,植物群落改造著周圍的環境,具有巨大的環境效應。在維持空氣的氧與二氧化碳平衡、凈化空氣、涵養水源、保持水土、防風固沙、調節氣候、改良土壤等方面,都具有重要作用。







{kind=link}