植物群落
出自 MBA智库百科(https://wiki.mbalib.com/)
植物群落(Plant Community)
目录 |
什么是植物群落[1]
植物群落是指在一定地段内,具有一定的植物种类组成、外貌和空间结构,各种植物之间及植物与环境之间彼此影响、相互作用的植物群体组合单元。一个地区全部植物群落共同形成的植物覆被层构成植被,植物群落是植被的结构单元。
植物群落的特征[1]
植物群落具有以下基本特征。
1.具有一定群落外貌
群落外貌即群落的外部形态。由于组成群落的植物种类不同,一个群落中的植物个体,分别处于不同的高度和密度,群落反映到人们眼中的样子也不同。人们到野外观察的时候,很容易辨认出来,这是森林、那是草原,就是由于群落的外貌不同。植物群落的外貌通常由构成群落的植物的生活型组成所决定。
2.具有一定群落结构
植物群落是植被的结构单元,具有一系列的结构特点。例如,形态结构上,包括垂直结构与水平结构、成层性、种的分布格局等。
3.具有一定种类组成
每个植物群落都由一定的植物种类组成。种类组成是区分不同植物群落的首要特征。一个植物群落中种类成分的多少及每种个体的数量,是度量群落多样性的基础。
4.形成一定的群落内部环境
植物群落对其居住环境产生重大影响,并形成群落内部特有的环境。例如,森林中的环境与周围的裸地就有很大的不同,包括光照、温度、湿度与土壤等都经过了植物群落的改造。即使植物非常稀疏的荒漠群落,土壤等环境条件也有明显改变。
5.具有一定的动态变化特征
群落是生态系统中有生命的部分。生命的特征是不停地运动。群落也是如此,其运动形式包括波动和演替等。
6.可以对群落进行分类与排序
群落因种类组成、外貌与结构、生态环境、演替特点的不同,可以区分为不同的类型,进行群落的分类。把一个地区内调查的植物群落样地,按相似度排定各样地的位序,从而分析各样地之间及其与周围生境之间的相互关系,称为植物群落的排序。
植物群落的性质[1]
1.整体性说(有机体学派、准有机体学派)
在植物生态学发展的早期,美国生态学家克莱门茨(1916;1928)认为,植物群落类似一个有机体:其理论根据是:任何一个植物群落都要经历一个从先锋阶段到相对稳定的顶极阶段的演替过程,如果时间充足的话,森林区的一片沼泽最终会演替为森林植被。这个演替过程类似于一个有机体的生活史。因此,群落像一个有机体一样,有诞生、生长、成熟和死亡的不同发育阶段,就像一个有机体的不同发育时期。他认为这种比拟是特别真实的,每一个顶极群落破坏后都能够通过基本上是同样形式的发展阶段再到达顶极阶段。
著名的法瑞学派创始人Braun-Blanquet(1928;1932)和另外一些学者(Nichols,1917;Warming,1909)把植物群落比拟为物种,把植物群落的分类看作与生物有机体的分类相似。正如物种是生物分类的基本单位一样,植物群落的最基本分类单位(植物群丛)则是植被的基本单位。在这种比拟中,他忽略了物种是一个遗传单位,而植物群落却在遗传上与其他相似的群落无关。群落间的从属关系不同于有机体之间的关系,它们仅是建立在某种结构和组成特征相似的基础上。
英国生态学家坦斯利(1920)虽然认为克莱门茨的有机体观点不正确,并指出在植物群落中,有些种群是独立的,它们在别的群落中也能很好地生长发育,相反,有些种群却具有强烈的依附性,即只能在这样的群落中而不能在别的群落中生长,群落中一些种群和某类群落并非严密结合,但他仍然主张群落具有整体性,应作为整体来研究,并用准有机体字样来形容群落,在此基础上他提出了生态系统概念,强调各个组成成分之间存在紧密联系。
以上学者的看法虽然有一定的差别,但都认为,植物群落是一个整体,从整体水平对外界发生反应。在某种程度上,类似于一个有机体,有自己的发生、发展与演变过程,各个群落间存在着明显的边界,植物群落可以和植物种一样进行分类与命名。每个植物群落是一个整体,而植被由不同的群落组成,所以植被具有不连续性。对群落这样的看法被称为整体性说。包括两个方面的基本认识:①群落的整体性(有机性或准有机性);②植被的不连续性。
2.个别性说(连续性观点、个体学派)
在整体性说流行时,H.A.Gleason经过审慎研究后,在1926年发表了“植物群丛中的个体论概念(individualistic concept)”一文,提出了恰好相反的观点,认为任何有关群落与有机体比拟都是欠妥的。因为群落的存在依赖于特定的生境与物种的选择性,但环境条件在空间与时间上都是不断变化的,因此群落之间不具有明显的边界。他指出长距离内连续观察的森林群落中,各物种及其多度的更替变化是渐变的,即强调植物群落内物种的个别性(或独立性)。即使在环境相对同质的地区,仍然可以见到植被的连续性变化,因而不存在群落的边界,而且在自然界没有任何两个群落是相同或相互密切关联的。传统群落分类实属人为产物。由于环境变化而引起的群落差异性是连续的。原苏联的R.G.Ramensky和美国的R.H.Whittaker均持类似观点。美国学者Whittaker(1970)的工作结果表明,在绝大多数情况下,不同种群之间的竞争并不产生明显的边界,也并不产生分布相似的种的集群。种群的中心和边界是沿着环境梯度散布的。这种观察结果与Gleason和Ramensky(1930)所主张的“种的个别性原则”相符合。他还认为,没有两个种分布相同,因为每个种都按照本身遗传的、生理的和生活周期特性,以及与环境条件和其他种的相互作用方式,以各自的方式分布。并且,由于不同种群沿着一个梯度分布,其生态位的重叠是疏松的,其中心是分散的,因而大多数群落总是沿着环境梯度连续地相互依次排列,而不形成明显的、可清楚划分的界限,植被存在着连续性。
强调种的独立性,认为植物是以种群(物种)为单位对环境作出反应,不存在明显的、可清楚划分界限的群落整体,在连续变化的环境梯度条件下,植被具有连续性,这样的观点称为个体学派。个体学派包括以下两点基本认识:①种的独立性;②植被的连续性。
根据这种看法,在研究植物群落时,更多地重视种群特征分析,重视随环境梯度的变化,种群性质出现的变化。强调种的独立性。认为植物群落可以出于某种需要进行分类,但其边界是主观人为划定的,没有客观性。他们用梯度分析与排序等定量方法研究植被,证明群落并不是一个个分离的、有明显边界的实体,多数情况下是在空间和时间上连续的一个系列。
植物群落的分类[1]
一、植物群落分类的原则
按照传统的整体学派观点,可以对植物群落进行分类。不同国家植被条件不同,植被研究中,对植物群落分类采用的分类标准也有所差异。目前我国采用的是“植物群落学~生态学原则”,它是以植物群落本身特征作为分类依据,同时又考虑到群落的生态关系。分类依据包括以下几方面。
(1)植物种类组成
种类组成是群落的最主要特征。群落的其他特征几乎全由这一特征决定。因此,在进行植物群落分类时考虑群落的种类组成是很自然的。中国植被主要采用优势种原则。把群落中各个层或层片中数量最多、盖度最大、对群落作用最明显的种,称为优势种。在主要层片(优势层片)中的优势种称建群种。在植被类型复杂,采用优势种原则有困难时,则可考虑采用生态幅度狭窄,对该类型有标志作用的标志种作为划分类型的标准。
(2)外貌和结构
外貌和结构是植物群落分类的传统依据。对未经充分研究的宏观区域的植被进行分类时,采用外貌和结构原则常可得到比较满意的结果。外貌主要根据生活型来划分。例如,按照生长型分类系统,从形态学的角度,把植物分为乔木、灌木、草本和叶状体等类群,以下再按形态和发育节律(落叶、常绿)划分第二级、第三级等。
(3)生态地理特征
由于历史原因,有时生活型和外貌不一定完全反映现代环境条件,按外貌原则划分的植被类型,常常包括了异质的类群。例如,针叶林在生活型和外貌上是相似的,但却分布在不同的生境里。在分类时,就应该考虑生态地理的因素,根据分布地区气候条件的不同,划分为寒温性针叶林、温性针叶林、暖温性针叶林、热性针叶林等。
(4)动态特征
优势种的原则着重于群落的现状,没有特别分出原生类型和次生类型。但是在某些情况下,需要考虑群落的动态演替特征。例如,我国森林区荒山坡灌草丛,按生态外貌原则应该是一独立的植被类型,但考虑到它的次生性质及与灌丛的演替关系,而把它与灌丛合为同一植被型组。
二、植物群落的分类系统
不同国家、不同地区的植被有各自的特点,不同学派考虑到所在地区的植被特征,往往采用不同的分类原则与依据。因此,形成了不同的植物群落分类系统,代表性的如英美学派、北欧学派、法瑞学派、苏联学派等。中国植被分类系统受苏联学派的影响较大。在《中国植被》(1980)一书中的系统采用植被型、群系、群丛3个基本分类单位,在各基本分类单位之上,各设一辅助单位,即植被型组、群系组与群丛组。并根据需要,在主分类单位之下设立亚级辅助单位,如植被亚型、亚群系等,以作为该级分类单位的补充,这样,得到九级分类单位,即植被型组、植被型、植被亚型、群系组、群系、亚群系、群丛组、群丛、亚群丛。
1.植被型组
植被型组是最高级的分类单位。凡建群种生活型相似,群落外貌、形态相似的植物群落联合为植被型组。我国1980年版《中国植被》共分了10个植被型组:针叶林、阔叶林、灌丛和灌草丛、草原和稀树干草原、荒漠、冻原、高山稀疏植被、草甸、沼泽、水生植被。它们主要是根据群落的外貌进行划分的。
2.植被型
植被型是最重要的高级分类单位,是在植被型组内,建群种生活型一级、二级相同或相近,同时对水热条件生态关系一致的植物群落联合。我国植被共分为29个植被型。例如,在针叶林植被型组下有5个植被型:寒温性针叶林,如兴安落叶松林;温性针叶林,如华山松林、油松林;温陛针阔叶混交林;暖性针叶林,如马尾松林;热性针叶林,如台湾松林。
建群种生活型相同或近似,反映了群落进化过程中对环境条件适应途径的一致,即其生态幅度和适应范围一致。就地带性植被而言,植被型是一定气候区域的产物,就非地带性植被而言,它是一定的特殊生境的产物。据此确定的植被型,大致有相似的结构、相似的生态性质,以及相似的发生和发展历史。
3.植被亚型
植被亚型为植被型的辅助单位,在植被型内根据优势层片或指示层片的差异进一步划分亚型。这种层片结构的差异,一般是由气候亚带的差异或一定的地貌、基质条件差异引起的。例如,寒温性针叶林植被型下划分两个亚型:寒温性落叶针叶林、寒温性常绿针叶林。雨林植被型(属于阔叶林植被型组)下划分为3个亚型:湿润雨林、季节雨林、山地雨林。
4.群系组
群系组是在植被型或植被亚型范围内,根据建群种亲缘关系近似(同属或相近属)、生活型(三级或四级)近似或生境相近而划分为群系组。划入为同一群系组的各群系,其生态特点一定是相似的。例如,典型草原可分出丛生禾草草原、根茎禾草草原和小半灌木草原群系组;温陛常绿针叶林(植被亚型)可以分出温陛松林、侧柏林、铁杉林等群系组。
5.群系
群系是最重要的中级分类单位。凡是建群种或共建种相同的植物群落联合为群系,以建群种命名,如马尾松林群系、兴安落叶松林群系、羊草草原群系。由于建群种或共建种相同,一个群系的结构、区系组成、生物生产力及动态特点都是相似的,因此,采用建群种(共建种)这一指标,就概括了群落各方面的特征。一般情况下,地带性群系主要分布在气候亚带范围内,非地带性群系则局限在某一特定生态因子的一定梯度范围内。在类型等级上,群系还常局限在某一植被亚型内。但对少数广生态幅的建群种,也会遇到例外的情况。例如,马尾松林,从北亚热带一直分布到南亚热带,同时在几个气候亚带内出现。又如,羊草草原和沟叶羊茅草原,在草甸草原和典型草原内均可遇到。还有极个别的种,如芦苇,不但在地域上分布很广,而且可形成沼泽、盐生草甸等不同植被类型,但在不同的生境中,其体态、群落外貌和结构均有很大变异。因此,按其最适生境将芦苇放入沼泽生境。
6.亚群系
亚群系是群系的辅助单位。在生态幅度比较广的群系内,根据次优势层片及其所反应的生境条件的差异而划分为亚群系。例如,羊草草原群系可划分出羊草一中生杂草、羊草一丛生禾草、羊草一盐中生杂草3个亚群系。但对大多数群系来说,并不需要划分亚群系。
7.群丛组
将层片结构相似,而且优势层片与次优势层片的优势种或共优种(某种情况下为标志种)相同的植物群落联合为群丛组,如羊草一丛生禾草亚群系之下的羊草一大针茅草原群丛组。
8.群丛
群丛是植被分类的主要基本单位。层片结构相同且各层片优势种或共优种相同的植物群落联合为群丛。换言之,属于同一群丛的植物群落,应具有相同的结构、每层具共同的优势种类、相同的生态特征、相同的动态特点、相似的生境、相同的季相变化并处于相同的演替阶段。命名:列出各层的优势种,不同层之间用“-”分隔,同层共优种之间用“+”分隔,如马尾松-杜鹃-铁芒萁群丛;马尾松+细叶青冈-杜鹃-铁芒萁+里白群丛。
植物群落的结构[1]
群落的结构,包括垂直结构和水平结构。
一、植物群落的垂直结构
群落的垂直结构,主要指群落的垂直分层现象,包括层与层片的分化。
1.层的分化
层(1ayer)的分化即不同的生物种出现在地面以上的不同高度和地面以下的不同深度,在整个群落内的垂直高度上出现许多层次的成层现象。
例如,森林植物群落根据组成植物高度的不同,地上部分一般可分为乔木层、灌木层、草本层和地被层4层。由于乔木层扎根很深,灌木及草本扎根较浅,因此,在群落的地下也有成层分化,这种分化和地上的层次分化是相对应的。
这种成层结构使植物充分利用环境中的空间:阳光、水分和矿质营养,扩大了植物利用环境的范围和效率。例如,在发育成熟的森林中,上层乔木可以充分利用阳光,穿过乔木层的光,有时仅占到达树冠的全光照的1/10,但林下灌木层却能利用这些微弱的、并且光谱组成已被改变了的光。在灌木层下的草本层能够利用更微弱的光,草本层往下还有更耐阴的苔藓层。
群落对光能的利用效率可以用叶面积指数来衡量。叶面积指数(LAI)是指单位土地面积上植物叶片的单面总面积。LAI=总叶片表面积/单位土地面积。
一些主要的天然植被类型,其叶面积指数如表1所示,由表1可知,叶面积指数与该群落的光能利用率有直接关系。叶面积指数越大,叶片交错重叠程度越大。群落对有效光能的利用效率越高。
表1 主要植被类型的叶面积指数与光能利用率(引自Barbour,et a1.1980)
| 植被类型 | LAI | 光能利用率% |
|---|---|---|
| 热带雨林 | 10-11 | 1.5 |
| 落叶阔叶林 | 5-8 | 1.0 |
| 北方针叶林 | 9-11 | 0.75 |
| 草原 | 5-8 | 0.50 |
| 冻原 | 1-2 | 0.25 |
| 草原化荒漠 | 1 | 0.04 |
| 农作物 | 3-5 | 0.60 |
注:光能利用率计算方法:单位面积净生产量(每克物质17.6kJ计)与单位面积全年接收的有效光合辐射(kJ)之比。
森林中的藤本和附生、寄生植物,它们攀援或附着在直立植株的不同高度,往往在整个群落的垂直高度内都有分布,因而并不形成一个层次,称为层间植物(interstratumplant)或层外植物(extrastratum plant)。
2.层片的分化
层片(synusia)词系瑞典植物学家Gains(1918)首创。他将层片划分为3级:第一级层片是同种个体的组合,第二级层片是同一生活型的不同植物的组合,第三级层片是不同生活型的不同种类植物的组合,即H.Gams的第一级层片指的是种群,第三级层片指的是植物群落。现在群落学研究中一般使用的层片概念,相当于Gams(1918)的第二级层片,即每一个层片均由同一生活型的不同植物所构成。因此,与层的分化只考虑高度不同,层片的分化主要考虑的是植物的生活型。
层片是植物群落结构的一个基本单位,由相同生活型或相似生态要求的种组成,同一层的植物可能属于不同的层片。例如,针阔叶混交林的乔木层,在层上可能是一层,但在二级层片上,包括了针叶层片和落叶阔叶层片两个层片。同一层片的植物也可能属于不同的层。例如,高位芽植物作为一个层片,植物的高度各不相同,可能属于乔木层或灌木层。
二、植物群落的水平结构
植物群落水平结构的主要特点是种间镶嵌性,即在植物群落内,种群与种群间分布的水平结构特征主要表现为“镶嵌性”,即同种植物常呈斑块状聚生在一起,不同物种的斑块镶嵌存在。例如,林冠下层光照强度强的林窗处,聚生着阳生灌木及草本,而光照强度弱的地方,只能生长耐阴植物。镶嵌性主要是由“生态因子的不均匀性”造成的,如小地形的差异、光照的强弱、土壤性质的不同及植物本身的繁殖特性等。
植物群落的种类组成[1]
种类组成是决定群落性质最重要的因素,也是鉴别不同群落类型的基本特征。怎样知道一个群落含有哪些物种及物种数量的多少呢?对于植物群落来说,通常采用野外实地调查的方法。每类群落所占的空间和位置不同,或大或小,或连续或分散。一般情况下不可能,也不需要把所有地段全面地进行清查,特别是数量特征更不可能如数查清。所以一般采取抽样调查方法完成这一任务。它的实质是选择调查有代表性的一定数量的小面积地段进行详细调查,以此估计推断此类群落整体。这些小面积地段称为样地。每块样地应该设在同样生境下同一类型植物群落中。
1.样地的形状
大多采用方形(长方形),故又称样方。也有使用圆形样地的,称为样圆。因为样圆很难划出边界,不及样方设置便利,故只在少数情况下使用小样圆调查,如对草原群落或森林下层草本层调查时可采用。长方形样地的长轴以平行于等高线为好,否则高差过大,样地内可能出现生境的变化,不好观察群落特征。样方的缺点是周边长,地处过界的植物增多,取舍不便;直角常测不准,出现闭合差,有时影响调查精度。调查林木蓄积量和草地产草量时要注意这类误差值。
小型样方(1m2)用于调查草本群落或者林下草本植物层。大型样方用于乔木群落或结构稀疏的群落。森林调查中样方常沿着预定的测线方向呈菱形设置。由中心点向前后定距离拉出样方对角线长度的4个点,然后从这4个点分别拉直长度恰为样方边长的测绳,使其在每一侧都恰好交接,就是样方的边界,这样可以避免确定直角的麻烦和误差。
如果山地坡度陡峻,按上述方法所设样地的投影面积将与实际面积有误差,须进行坡度订正,表2中列有在各种坡度上应采用的一半对角线的长度值。
2.样地大小
群落的最小面积是指包含群落中大多数植物种类,并能表现群落主要结构特征的最小地段面积,它是进行群落野外调查的最小面积。找出群落最小面积一般使用种一面积曲线法,其方法是:从较小面积开始统计植物种类数目,然后逐次向外围扩大,同时登记新发现的植物种类,直到基本不再增加新种类为止,最后绘制植物种数的面积曲线:以面积大小为横轴,以种数为纵轴,填入逐次调查数值,并连成平滑曲线,在曲线由陡变缓转折相对应的面积就是群落最小面积(图1)。
下列样地面积的经验值可供参考使用。
草本群落:1~1Om2
灌丛:16~1OOm2
单纯针叶林:1OOm2(10m×1Om)
复层针叶林、夏绿阔叶林:400~500m2(20m×20m,20m×25m)
亚热带常绿阔叶林:800~1200m2
热带雨林和山地雨林:2500~4000m2
通过各种群落类型的对比,可以发现,环境条件越优越,组成群落的植物种类就越多,群落的结构就越复杂,群落的最小面积相应地就越大。例如,我国西双版纳南部的热带雨林群落,它的最小面积至少为2500m2,其中包含了组成群落的主要高等植物130种左右;而东北小兴安岭红松林的一个群落中,最小面积约为400m2,包含主要的高等植物40种左右。
3.样地数目
最小面积是群落结构的一项特征,也是设定样地面积的依据。但一块“最小面积”的样地并不能包含一个群落的全部植物种类,更不要说所属群落类型的全部植物种类了。确定一个群落的特征,需要有多个群落样地资料,否则就不完整。
样地数目的多少,取决于群落结构的复杂程度。如果群落内部分布和结构都较均一,少数样地就能很好地表现出群落特征。当群落的结构复杂并且变化较大,植物分布不规则时,必须增加样地数目才能提高调查资料的可靠性。但调查样地是要付出人力、物力的,所以要考虑经济效益。究竟取多少样方,这决定于研究的精度要求。根据统计检验理论,多于30个数值(大样本)才较可靠。
此外,所有样地必须依照顺序编号,或再加以分类排号,这样做能避免混乱,整理资料时用代号也很方便。
4.样地的布局
一块样地的面积可能仅占某类群落在一个地区总面积几百分之一或几千分之一,各个样地在这一范围内应当怎样布局也很重要,它影响着调查结果的准确程度。
(1)主观(有意识)取样
就是从群落所在空间中,主观地选择被认为有代表性的(典型的)或者有某种特点的地块作为调查样地,这在路线式粗放调查中常常采用。所选样地要注意环境条件的一致性与群落外貌的一致性,最好处于群落的中心部位,避免过渡地段。但这种数据容易有偏差或遗漏,特别是一些不显眼的、较小或分散的特征可能被忽视,因此,这种数量资料不能用于统计分析。
(2)系统取样(机械取样)
就是严格按照一定的规则(方向和距离)确定样地位置。在面积十分广阔的森林和草地中这是一种通用方法。它的优点是布点均匀,寻址简便。缺点是当群落的某些特征呈不规则的随机分布时,就可能使某些特征不能如实表露;或者某些呈规律分布的特征(如沙丘和丘间洼地)恰好和样地分布规律不一致,其结果可能就不能反映规律性。
(3)随机抽样
这种抽样的基本原则是,使某类群落各个部分都有同等机会(等概率)被抽取做样地。为此,应当把要调查的地段分成大小均匀的若干部分,每部分都编号或确定坐标位置,利用随机数字表、掷骰、抽签等方法随机选出一定数量的、占有一定位置的样地。地块的选择可直接在现场进行,也可选在地图上进行,然后在现场落实。此法优点是,调查的数据可以应用于各种统计处理,进行可靠性检验。许多人认为这是唯一科学有效的方法,但缺点是它常需要较多的样地,工作量较大,也有个别的种类易被遗漏。
(4)部分随机取样(分层抽样)
部分随机取样为系统取样和随机取样的混合法。先将群落地段系统均匀划分好地段(编号),每个分地段中再随机抽取样地。这使所研究的地段内各点被抽取的机会加大了,资料也能用于统计,但是工作量仍然较大。
经过样地调查,可以发现,有一些群落,所含有的物种数量较多,有些则较少。有些群落的结构较为复杂,有些较为简单。
一般来说,环境条件越优越,群落的结构越复杂,组成群落的生物种类越多;环境条件越恶劣,群落的结构越简单,组成群落的生物种类就越少。
群落的复杂程度和物种多少可以用物种多样性来表述。
物种多样性(species diversity)是生物多样性的一个方面,是指一个群落中物种数目的多少及物种个体数目分布的均匀程度,表示物种的丰富度及其可持续程度。包括两个方面的内容。
1)种的丰富度(richness of species):物种数量的多少。
2)物种分布的均匀程度(eveness of species distribution):个体数量在种间分布的均匀程度。
物种多样性表达的是物种丰富度及其可维持程度。例如,假设共有100个个体,有A和B两个物种,甲地有A与B各50个个体,乙地A有99个个体,B有1个个体,则甲地的可维持度就远高于乙地。因为乙地B物种的1个个体如果死亡,则乙地就只剩A这1个物种了。
物种多样性指数表示方法很多。以Simpson(辛普生)多样性指数和Shannon-Wiener指数(香农一威纳指数)最为常用。
辛普生多样性指数(Simpson’s diversity index)的生物学意义是:在很大的群落中,随机抽取两个个体,它们为不同物种的概率是多少?Simpson于1949年提出:在不限大小的群落中,随机抽取得到两个个体,它们是同一物种的概率与物种多样性有关。例如,在加拿大北部寒带森林中,随机采取两株树它们属于同一个种的概率就很高,相反,如在热带雨林中随机取样,两株树是同一种的概率很低。他从这个想法出发得出多样性指数。
辛普生多样性指数=随机取样的两个个体属于不同种的概率
=1-随机取样的两个个体属于同种的概率
设总个体数为N,第i个物种的个体数为ni,Pi = ni / N(Pi为相对多度;除此之外,也可用相对优势度、相对盖度等)。辛普生多样性指数D的计算公式为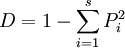
例如:总个体数为N,第i个物种的个体数为ni,Pi = ni / N;
假设N=1000,共2个物种,ni = 500,n2 = 500,则:
P1 = 1 / 2,P2 = 1 / 2,L = 1 − [(P1)2 + (P2)2] = 0.5。
假设ni = 1,ni = 990,则:P1 = 1 / 1000,P2 = 999 / 1000,L = 1 − [(P1)2 + (P2)2] = 0。
辛普生多样性指数的生物学意义表示的是在群落中任意选取两个个体,这两个个体是不同物种的概率。其值大小为0-1。辛普生指数值越大,则表示该群落的物种多样性越大。
Shannon-Wiener指数(香农-威纳指数)的计算公式为
式中,H为Shannon-Wiener指数,与Simpson指数相同;Pi = ni / N,Pi为第i种的相对多度;Ni为第i种的个体数;N为群落中的个体总数;S为物种种数。N也可以采用其他数量参数,如重要值、密度、频度等。Shannon-Wiener指数来源于信息理论。它的计算公式表明,群落中生物种类增多代表了群落的复杂程度增高,即H值越大,群落所含的信息量越大,群落的物种多样性越大。
群落物种多样性越大,则含有的生物基因越丰富,食物链越复杂,群落的自我调节能力就越强,生态平衡就越不容易遭到破坏,这叫做物种多样性导致群落稳定性原则。
此外,在群落交错区(ecotone),即两个或多个群落之间的过渡地带,如海陆交界的潮间带、河口湾、森林与草原等交界地带的群落中,生物种类和数量常比单一类型群落中的多,物种多样性指数高,这种现象称为边缘效应(edge effect)。
群落交错区,又称生态交错区或生态过渡带,如森林和草原之间的森林草原地带,两个不同的森林类型之间或两个草本群落之间的地带等。这种过渡带有的宽、有的窄;有的是逐渐变化,有的突然变化。群落交错区是一个交叉地带,也是一个种群竞争的紧张地带,往往包括包含两个重叠群落中的一些种及交错区本身所特有的种,这是因为群落交错区的环境条件比较复杂,能为不同生态类型的植物定居,并且为更多的动物提供食物、营巢和隐蔽条件。例如,我国大兴安岭森林边缘,每平方米植物达30种以上,明显高于其内侧的森林群落与外侧的草原群落。
群落成员型是根据物种在群落中的作用和地位而划分的物种类群。假设常绿阔叶林样方,400m2以内,乔木层内大头茶共100株,栲树10株,马尾松3株,那么明显可以看出,同样是该群落内的组成物种,大头茶在群落中所占地位最重要,其他物种则较次要。因此,可以区分出优势种、伴生种等不同的类型。
优势种:群落的优势种类,是指群落每层中个体数量大、生物量大、覆盖程度大的生物种类。
建群种群落的主要层次(优势层)的优势种称为建群种,它决定着群落的内部结构和特殊环境条件。
共优种:群落各层的优势种如果不止一种,则称为共优种。
共建种:如果群落中有不止一个的建群种则称为共建种,即在优势层中有两个以上的种占优势。
如果群落中的建群种只有一个,称为“单优种群落”。如果具有两个或两个以上同等重要的建群种,就称为“共优种群落”或“共建种群落”。热带森林几乎全是共建种群落,北方森林和草原则多为单优种群落,但有时也存在共优种,如由贝加尔针茅和羊草共建的草甸植物群落。
生态学上的优势种对整个群落具有控制性影响。如果把优势种去除,必然导致群落性质和环境的变化;但若把非优势种去除,只会发生较小的或不显著的变化。因此,不仅要保护那些珍稀濒危植物,而且也要保护那些建群植物和优势植物,它们对生态系统的稳定起着举足轻重的作用。
附属种:这类植物虽然在群落中出现,参加到群落的组成中去,但对群落内环境所发生的作用不及优势种,即群落中除优势种以外的其他生物种类。
附属种中,根据作用不同,又区分为以下几种。
亚优势种:个体数量和作用都次于优势种,但在决定群落性质和控制群落环境方面仍起着一定作用的植物种。
伴生种:群落中相当稳定的与优势种伴生在一起的种类。虽然在群落中出现,但对群落内环境所起的作用不及优势种和亚优势种。
标志种:生态幅度狭窄对群落有标志作用的物种。
偶遇种:在群落中出现频率很低的种类。可能是由于外界因素(如动物、人类作用)的影响偶然进入群落的物种,也可能是衰退中的残遗种。
同一个植物种在不同的群落中可以以不同的群落成员型出现。例如,在内蒙古高原中部排水良好的壤质栗钙土上,针茅是建群种,羊草则是亚优势种或伴生种;但在地形略为低凹,有地表径流补给的地方,羊草则是建群种,针茅退居其次。而当强度放牧时,小半灌木冷蒿成为建群种,羊草和针茅成为次要成分。
1.多度
多度(abundance)表示一个种在群落中的个体数量。植物群落中植物种间的个体数量对比关系,可以通过各个种的多度来确定。多度的统计方法,通常有两种:一种是个体的直接计算法,即“记名计算法”,即在一定面积的样地中,直接点数各种群的个体数目。在森林群落对乔木层的调查中,或者详细的群落研究中常用;另一种是目测估计法。一般在植物个体数量多而植物体形小的群落(如灌木、草本群落),或者在踏察中常用。目测估计法是按预先确定的多度等级来估计单位面积上个体的多少。
常用的多度等级是德氏(Drude)的七级多度制:Soc.极多;Cop3.很多;Cop2.多;Cop1.尚多;Sp.少;S01.稀少;Un.单一个体。
相对多度(relative abundance):样地内某一种的个体数占全部种个体数的百分比。
RA=N(某个种个体数)/∑N(全部种个体数之和)×100%
2.密度
密度(density)是指单位面积上的植物株数,用公式表示为
d=N/S
式中,d为密度;N为样地内某种植物的个体数目;S为样地面积。
密度的倒数即每株植物所占的单位面积。
样地内某一种的密度占全部种密度总和的百分比称为相对密度(relative density)。相对密度:RD=d/∑d×100%。在计算上,等于相对多度。
3.盖度
盖度分为以下几种。
投影盖度:植物地上部分垂直投影所覆盖的面积,简称盖度。
郁闭度:森林上层总投影覆盖地面的比率。大小值用0~1来表示。0代表覆盖度为0,1代表覆盖度为100%。
基盖度:植物基部覆盖面积。乔木通常用离地1.3m高处的胸高断面积作为基盖度。草本以离地面1英寸(2.54cm)高度的断面积计算。
显著度(prominance):样地中一个乔木种的所有个体基盖度之和,也称优势度(dominant)。
相对显著度(relative pominance):群落中某一物种的显著度占所有种显著度之和的百分比。RP=P/∑P×100%
4.频度
频度(frequency)是指某物种在调查样地内出现的频率。
频度一某物种出现的样地数/样地总数×100%
相对频度:某一物种的频度与全部物种频度之和的比。
RF=F(某个种频度)/∑F(全部种频度之和)×100%
丹麦学者拉恩基尔(C.Raunkiaer)用0.1m2的小样圆任意投掷,将小样圆内的所有植物种类加以记载,就得到每个小样圆的植物名录,然后计算每种植物出现的次数与样圆总数之比,得到欧洲草地群落中各个种的频度,并根据8000多种植物的频度统计结果,编制出标准频度图解。通过样圆对草原植物群落进行数量统计,是一种方便、常用的方法。
5.重要值
重要值(important value)是美国威斯康星(Wisconsin)学派提出的用来表示某个种在群落中地位和作用的综合数量指标。因为它简单、明确,所以得到了广泛应用。
主要用于森林群落乔木层个体重要值的计算。
重要值=相对密度(%)+相对频度(%)+相对显著度(%)
IV=RD%+RF%+RP%
结果值常再除以300或3。重要值最大的种即为优势种。
6.高度
高度为测量植物体体长的一个指标。测量植物取其自然高度或绝对高度:高度分为最大高度、最小高度、平均高度、枝下高度等,要分别测定。在森林群落中,可以使用测高仪测定乔木的高度。
7.体积
体积是生物所占空间大小的度量。在森林植被研究中,这一指标特别重要。在森林经营中,通过体积的计算可以获得木材生产量(材积)信息。木材的体积称为材积,可以通过下式计算:材积=胸高断面积×树高×形数
形数是树干体积与等高同底的圆柱体体积之比。这是由于树木体形是圆锥形而不是圆柱形,而圆锥形的尖削度又因各个树种不同,因此在断面积乘树高获得圆柱体体积之后,必须按不同树种乘以该树种的形数(在制就的森林调查表中查得),才能获得乔木的体积。草本植物或小灌木体积的测定,可用排水法进行。
8.重量
重量(质量)是用来衡量植物种群生物量(biomass)或现存量(standing crop)多少的指标。生物量指某一时刻单位面积内实际存活的有机物质总量,通常用kg/m2或t/hm2表示。广义的生物量是生物在某一特定时刻单位空间的个体数、重量或其含的能量,可用于指某种群、某类群或整个生物群落的生物量。狭义的生物量仅指重量,可以是鲜重或干重。某一特定时刻的生物量称为现存量(standing crop)。生产力则是某一时间段内由活的生物体新生产出的有机物质总量。在草原植被研究中,重量指标特别重要。单位面积或容积内某一物种的重量占全部物种总重量的百分比称为相对重量。
植物群落的内部环境[1]
环境既是形成植物群落的要素,也是群落存在的条件。反过来,群落的存在又必然对环境起到影响,在一定程度上改变环境。一方面,群落的组成、结构、功能、成因、动态、分布无不受到环境条件的制约;另一方面,群落生活也影响着环境,决定着环境的许多特性,并创造着群落的内部环境。在植物群落内部,由于群落本身的作用而对环境产生影响,在群落内会形成特殊的生境,称为植物群落的内部环境。以森林为例,在森林植物群落内,与森林外相比,生境有如下变化:弱光、并且主要为漫射光;弱风至静风;温差小;湿度大;土壤腐殖质多;氧气浓度高,CO2浓度较小;且CO2浓度一般从近地面处至林冠逐渐减少等。
一、群落内的光照
在群落内部,由于植物群落对光辐射的吸收、反射和透射作用,所以无论是在森林还是草本群落的植冠下,入射的光量总比其他条件相同而没有植被的陆地表面少得多。
穿过枝叶间隙射入群落内部的直射光,形成大小不等的光斑,虽然其成分改变很少,但照射强度显著减弱。
通过群落上层进入林下的光,由于群落对光的“过滤”作用,光的成分和强度都发生了变化。由于大部分有效辐射被叶片吸收,林下以散射光为主,强度上也显著降低。
可使用光照度计,通过测定相对光照度来定量光减弱的程度。
二、群落内的温度
群落内与群落外的温度相比波动较为缓和。一般群落内的平均温度白天和夏天低于群落外空旷地面,而夜间和冬季则高于群落外空旷地面。昼夜和全年的温度变幅较小。
群落的结构越复杂,郁闭度越高,对温度的作用越显著。大面积的森林,群落对温度的作用不仅在林内,还可以影响一定范围的地方气候。
三、群落内的水分
植物群落能截留降水,保蓄水分。
如在河流的上游营造大面积森林,可以起到涵养水源的作用;在山坡地带建造森林,可以减少地面径流,防止水土流失。因此,植物群落在涵养水源、保持水土方面有重要意义。
降落到群落的水分,一部分到达地面,一部分被树冠截留。群落截留水分的能力,一与群落结构、郁闭度有关;二与降水强度有关,降水强度越小,截留越多;三与土壤的结构和性质有关。土壤结构差、质地粗,保水能力差;四与地形起伏大小有关,地形起伏大也不能有效地保留水分。
四、群落内的空气
植物群落对空气的改变主要表现在两个方面。
空气成分发生变化:由于植物的光合作用,群落内CO2浓度降低,O2浓度增大,尤其是靠近林冠上层,这种变化最为明显。
森林不但可以增加氧气,还可以吸附粉尘;对一些有毒气体如SO2、HF、Cl2等有一定吸收能力,是天然的空气净化器。
抑制风速:由于群落对气流的阻抑,群落内表现为弱风,在结构复杂郁闭度高的森林内,甚至表现为静风状态。
五、群落内的土壤
植物群落对土壤最明显的作用是可以增加土壤腐殖质含量,增强土壤肥力。
此外,植物群落在改善土壤理化性质、优化土壤结构、固定流沙、防止和治理盐碱化方面都有显著作用。
植被类型通常与一定的土壤条件密切关联,如热带雨林—砖红壤、北方针叶林—灰化土、温带草原—黑钙土或栗钙土等,二者共同受到气候条件的控制。
综上所述,植物群落改造着周围的环境,具有巨大的环境效应。在维持空气的氧与二氧化碳平衡、净化空气、涵养水源、保持水土、防风固沙、调节气候、改良土壤等方面,都具有重要作用。







{kind=link}